Acabamos de ver que las soluciones de los sistemas unidimensionales se comportan de forma bastante trivial. Pero la dependencia de los parámetros añade picante porque pueden producirse cambios cualitativos en el comportamiento de las soluciones. Estos cambios se llaman bifurcaciones.
El objetivo de este capítulo es presentar algunos ejemplos y no desarrollar la teoría completa. Exploraremos otros tipos de bifurcaciones cuando pasemos a sistemas bidimensionales y tridimensionales.
El escenario básico es una ecuación diferencial unidimensional $\dot{x}=f_\mu(x)$, donde $\mu$ es un parámetro. Esperamos que la variación de $\mu$ pueda causar la aparición o destrucción de equilibrios y también afectar a su estabilidad.
Un modelo de pesquería con bifurcación de nodos en silla de montar
La ecuación $\dot{x}=rx\left(1-\frac{x}{K}\right)-h$ proporciona un modelo demasiado simplificado de una pesquería. En ausencia de pesca, se supone que la población crece según la ecuación logística. Los efectos de la pesca se modelan mediante el término $-h$, que dice que los peces se capturan a un ritmo constante $h>0$, independientemente de su abundancia $x$. Esto supone que los pescadores capturan el mismo número de peces todos los días, sin preocuparse de agotar la población.
A priori, tenemos tres parámetros, pero el sistema puede reescribirse en forma adimensional como
$$ \dot{x}=x(1-x)-\mu $$
mediante un reescalado adecuado. Ya mencionamos la no dimensionalización cuando discutimos el modelo de Volterra para tiburones y sardinas en el primer capítulo. Aquí vemos la utilidad de este procedimiento, ya que comprobamos que, de hecho, el sistema sólo depende de un único parámetro (adimensional).
Podemos observar los siguientes comportamientos.
Los puntos fijos son las soluciones de la ecuación cuadrática $x(1-x)-\mu=0$. Hay dos equilibrios si $\mu\in (0,1/4)$, uno es estable y el otro inestable. Cuando $\mu=1/4$, estos equilibrios se unen en un punto fijo ‘semi-estable’ que corresponde a la solución única de la ecuación cuadrática. En cuanto $\mu>1/4$, este punto fijo desaparece y no hay ningún equilibrio.
Decimos que se ha producido una bifurcación en $\mu=\frac{1}{4}$. Esta bifurcación se llama una bifurcación de nodo de silla de montar. (La razón de ese nombre se hará evidente cuando veamos una bifurcación completamente análoga en dimensión dos). Es el mecanismo básico por el que se crean y destruyen puntos fijos: al variar un parámetro, dos puntos fijos se acercan entre sí, ‘chocan’, y se ‘aniquilan’ mutuamente.
Un ejemplo de bifurcación transcrítica
El modelo anterior tiene algo de tonto porque la población puede volverse negativa. Un modelo mejor tendría un punto fijo en la población cero para todos los valores del parámetro. La forma más sencilla de conseguirlo es fijar
$$ \dot{x}=x(1-x)-\mu x $$
donde $\mu>0$ representa la fracción de la población que los pescadores capturan por unidad de tiempo. Si $r=1-\mu$, esta ecuación se convierte en
$$ \dot{x}=rx-x^2. $$
Se parece a la ecuación logística pero, olvidándonos de su interpretación como modelo poblacional, ahora permitimos que $x$ y $r$ sean positivos o negativos. Independientemente del valor de $r$, $0$ es un punto fijo pero vamos a ver que cambia su estabilidad al variar $r$, como muestra el siguiente experimento.
Observamos que, para $r<0$, existe un punto fijo inestable $\bar{x}=r$ y el origen $0$ es estable. A medida que $r$ aumenta, el punto fijo inestable se aproxima al origen y coalesce con él cuando $r=0$. Finalmente, cuando $r>0$, el origen se vuelve inestable, y $\bar{x}=r$ es ahora (asintóticamente) estable. Podemos decir que se ha producido un intercambio de estabilidades entre los dos puntos fijos. Esta bifurcación se ha producido en $r=0$. El ejemplo que consideramos puede parecer particular, pero en realidad no lo es: cerca de una bifurcación transcrítica, la dinámica se parece a $\dot{x}=rx-x^2$.
Un modelo de brote de insectos
Terminamos este capítulo con un ejemplo biológico de bifurcación y catástrofe, a saber, un modelo de brote repentino de un insecto llamado gusano de la yema del abeto. Cuando se produce un brote, este insecto puede llegar a defoliar y matar el abeto balsámico en los bosques del este de Canadá en unos cuatro años. La versión más sencilla para un modelo de la interacción entre los gusanos de la yema y el bosque se basa en una separación de escalas temporales: la población de gusanos de la yema evoluciona a una escala temporal mucho más rápida que los árboles. Así, en una primera aproximación, las variables del bosque pueden tratarse como constantes. El modelo propuesto para la dinámica del gusano cogollero es el siguiente
$$ \dot{x}=\rho x\left( 1-\frac{x}{\kappa}\right)-\frac{\beta x^2}{\alpha^2+ x^2}, $$
donde los parámetros $\alpha,\beta,\kappa,\rho$ son parámetros positivos. En ausencia de depredadores, se supone que la densidad de población de gusanos cogolleros $x(t)$ crece logísticamente con una tasa de crecimiento $\rho$ y una capacidad de carga $\kappa$. La capacidad de carga depende de la cantidad de follaje que quede en los árboles, pero aquí la tratamos como fija, como ya se ha mencionado. El término $\frac{\beta x^2}{\alpha^2+ x^2}$ representa la tasa de mortalidad debida a la depredación, principalmente por aves.

La forma específica para la tasa de mortalidad tiene en cuenta, de la forma más simple, los siguientes comportamientos. Casi no hay depredación cuando los gusanos son escasos; los pájaros buscan comida en otra parte. Pero, cuando la densidad de población supera un cierto nivel crítico $x=\alpha$, la depredación se activa bruscamente y luego se satura.
Ahora cabe preguntarse: ¿qué se entiende por brote? Antes de responder a esta pregunta, conviene refundir el modelo en una forma adimensional, como hicimos antes para modelos más sencillos. En efecto, el modelo tiene cuatro parámetros (positivos): $\rho,\kappa,\alpha$ y $\beta$. Si te pones a jugar a reescalar la ecuación, verás que hay varias formas de hacerlo. Resulta, después de algún ensayo y error, que una opción muy conveniente es deshacerse de los parámetros en el término de depredación. Tras un poco de álgebra, se obtiene el modelo adimensional
$$ \dot{x}=rx\left(1-\frac{x}{K}\right) - \frac{x^2}{1+x^2}\, \cdot $$
Análisis de puntos fijos.
Siempre tenemos el punto fijo $x=0$ y no es difícil comprobar que siempre es inestable aplicando la prueba de la derivada. Intuitivamente, esto no es sorprendente ya que para $x$ muy pequeños, tenemos aproximadamente $\dot{x}\approx rx$. Los otros posibles puntos fijos de la ecuación vienen dados por las soluciones de la ecuación
$$ g(x)=h(x) $$
donde
$$ g(x)=r\left(1-\frac{x}{K}\right)\quad \text{and}\quad h(x)=\frac{x}{1+x^2}\, \cdot $$
Obsérvese que la función $h$ es independiente de los parámetros; sólo la función $g$, cuya gráfica es una recta, se desplaza al variar $r$ y $K$. Esto se debe a nuestra elección de la no dimensionalización.
Como muestran los experimentos, si $K$ es suficientemente pequeño, hay exactamente una intersección para cualquier $r>0$. Pero, para $K$ grandes, podemos tener una, dos o tres intersecciones, dependiendo del valor de $r$. Podemos observar que, cuando estamos en el intervalo de parámetros para el que hay tres intersecciones, a medida que disminuimos $r$ con $K$ fijo, los puntos fijos $b$ y $c$ se aproximan entre sí y acaban uniéndose. Esta es la firma de una bifurcación de nodo de silla de montar. Después de la bifurcación, sólo queda un punto fijo (además de $0$, por supuesto). Otra forma de variar los parámetros es fijar $K$ y, a partir de $r$ pequeño, observar que $a$ y $b$ pueden colisionar y aniquilarse al aumentar $r$.
Los experimentos también muestran cómo cambia la estabilidad de los puntos fijos al variar $r$ y $K$. Para los valores correspondientes a tres puntos fijos, el punto fijo estable más pequeño $b$ se denomina nivel de refugio de la población de gusanos cogolleros, mientras que el punto fijo estable más grande $c$ es el nivel de brote. Desde el punto de vista del control de plagas, se desea mantener la densidad de población en $a$ y lejos de $c$. El destino del sistema viene determinado por la condición inicial $x_0$. Se produce un brote si y sólo si $x_0>b$; el punto fijo inestable $b$ desempeña el papel de umbral.
Otro escenario más interesante que conduce a un brote es una bifurcación de nodo de silla de montar. En efecto, imaginemos que ahora $r$ y $K$ derivan de tal manera que el punto fijo $a$ desaparece; entonces la población saltará repentinamente al nivel de brote $c$. De hecho, la situación es aún peor porque existe el llamado efecto de histéresis: aunque los parámetros vuelvan a sus valores anteriores al brote, ¡la población no volverá a descender al nivel de refugio!
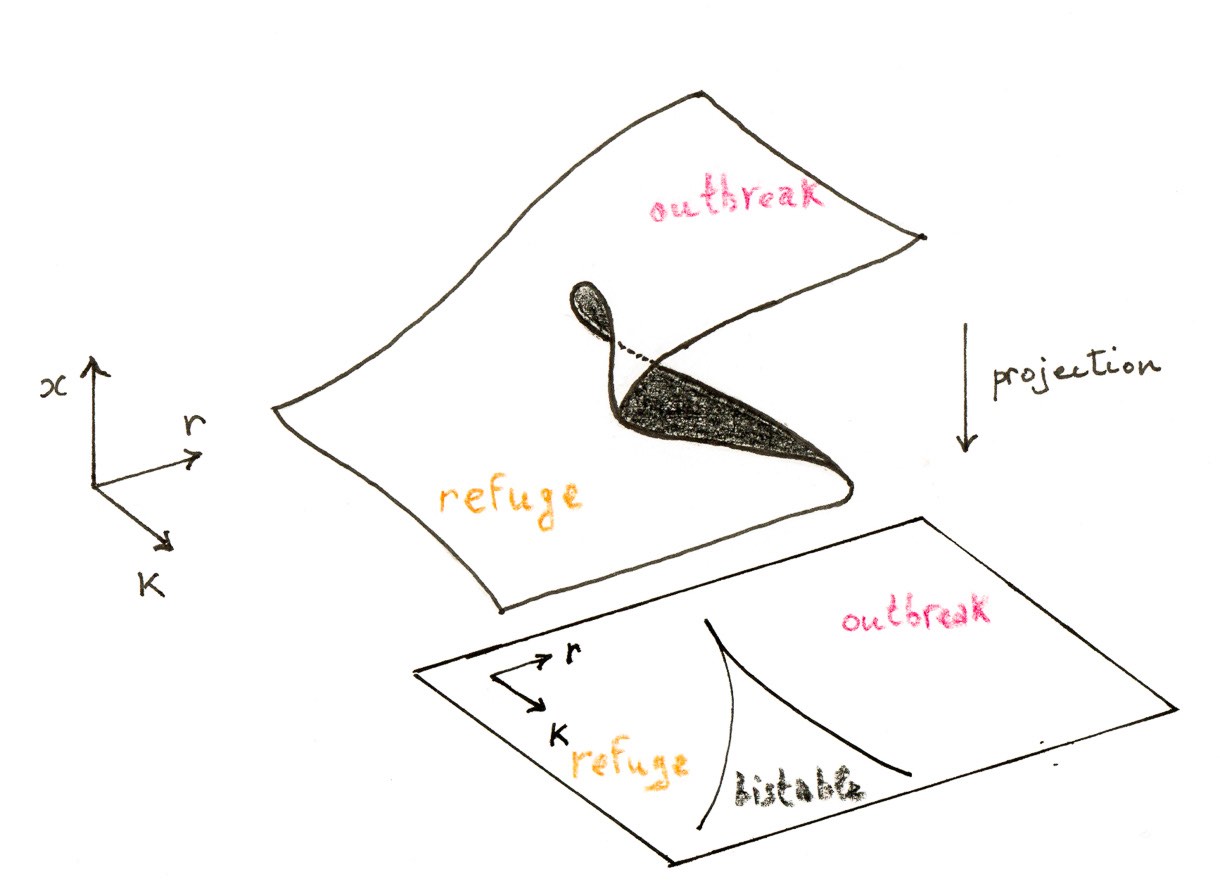
La riqueza de este sencillo modelo de insecto es bastante notable. Ilustra el hecho de que no debemos subestimar los modelos unidimensionales cuando se producen bifurcaciones.