Nous avons vu que les systèmes unidimensionnels présentent, à première vue, un comportement assez basique. Toutefois, la dépendance aux paramètres introduit une richesse nouvelle, en rendant possibles des modifications qualitatives du comportement des solutions.
Ce chapitre vise à introduire quelques exemples et non à développer la théorie complète. Nous verrons d’autres types de bifurcations lorsque nous passerons à des systèmes bidimensionnels et tridimensionnels.
Le cadre de base est une équation différentielle unidimensionnelle $\dot{x}=f_\mu(x)$, où $\mu\in\mathbb{R}$ est un paramètre. On s’attend à ce qu’une variation du paramètre $\mu$ puisse entraîner la création ou la disparition de points d’équilibre, tout en influençant leur stabilité.
Modèle d’une pêcherie avec une bifurcation col-nœud.
L’équation $\dot{x}=rx\left(1-\frac{x}{K}\right)-h$ fournit un modèle ultra simplifié d’une pêcherie. En l’absence de pêche, la population est supposée croître selon l’équation logistique. Les effets de la pêche sont modélisés par le terme $-h$qui indique que les poissons sont capturés à un taux constant $h>0$, indépendamment de leur abondance $x$. Cela suppose que les pêcheurs attrapent chaque jour le même nombre de poissons, sans se soucier d’épuiser la population.
A priori, nous avons trois paramètres, mais le système peut être réécrit sous forme adimensionnée :
$$ \dot{x}=x(1-x)-\mu $$
par une remise à l’échelle appropriée. Nous avons déjà mentionné la non-dimensionnalisation lorsque nous avons abordé le modèle de Volterra pour les requins et les sardines dans le premier chapitre. Nous voyons ici l’utilité de cette procédure puisque nous constatons qu’en fait, le système ne dépend que d’un seul paramètre (sans dimension).Nous pouvons observer les comportements suivants.
Les points fixes sont les solutions de l’équation quadratique $x(1-x)-\mu=0$. Il existe deux équilibres si $\mu\in (0,1/4)$, l’un est stable et l’autre est instable. Lorsque $\mu = 1/4$, ces points d’équilibre fusionnent en un point fixe « semi-stable » correspondant à l’unique solution de l’équation quadratique. Dès que $\mu > 1/4$, ce point fixe disparaît et il n’existe plus aucun équilibre.
On dit qu’une bifurcation s’est produite pour $\mu=\frac{1}{4}$. Cette bifurcation est appelée une bifurcation selle-nœud (ou nœud-col). (La raison de ce nom deviendra évidente lorsque nous verrons cette bifurcation en dimension deux). C’est le mécanisme de base par lequel les points fixes sont créés et détruits : à mesure qu’un paramètre varie, deux points fixes se rapprochent, « entrent en collision » et s’« annihilent » mutuellement.
Un exemple de bifurcation transcritique.
Le modèle précédent présente un défaut majeur : il permet à la population de devenir négative, ce qui n’a aucun sens sur le plan biologique. Un meilleur modèle aurait un point fixe à la population nulle pour toutes les valeurs du paramètre. La façon la plus simple d’y parvenir est de considérer
$$ \dot{x}=x(1-x)-\mu x $$
où $\mu>0$ représente la fraction de la population que les pêcheurs capturent par unité de temps. En fixant $r=1-\mu$, cette équation devient$$ \dot{x}=rx-x^2. $$
Cela ressemble à l’équation logistique mais, oubliant son interprétation en tant que modèle de population, nous permettons maintenant à $x$ et $r$ d’être positifs ou négatifs. Quelle que soit la valeur de $r$, $0$ est un point fixe mais nous allons voir qu’il change de stabilité quand on fait varier $r$, comme le montre l’expérience suivante.
On observe que, pour $r<0$, il existe un point fixe instable $\bar{x}=r$ et que l’origine $0$ est stable. Lorsque $r$ augmente, le point fixe instable se rapproche de l’origine et coïncide avec elle lorsque $r=0$. Enfin, lorsque $r>0$, l’origine devient instable, et $\bar{x}=r$ est maintenant (asymptotiquement) stable. On peut dire qu’un échange de stabilités a eu lieu entre les deux points fixes. Cette bifurcation s’est produite en $r=0$. L’exemple que nous considérons peut sembler particulier mais il ne l’est pas en fait : proche d’une bifurcation transcritique, la dynamique ressemble $\dot{x}=rx-x^2$ (en un sens qui peut être rendu rigoureux).
Un modèle d’épidémie d’insectes
Nous terminons ce chapitre par un exemple biologique de bifurcation et de catastrophe, à savoir un modèle d’épidémie soudaine d’un insecte appelé tordeuse des bourgeons de l’épinette. Lorsqu’une épidémie se produit, cet insecte peut effectivement défolier et tuer le sapin baumier dans les forêts de l’est du Canada en quatre ans environ. La version la plus simple d’un modèle d’interaction entre les tordeuses et la forêt repose sur une séparation des échelles de temps : la population de tordeuses évolue sur une échelle de temps beaucoup plus rapide que les arbres. Ainsi, en première approximation, les variables de la forêt peuvent être traitées comme des constantes. Le modèle proposé pour la dynamique de la tordeuse est le suivant :
$$ \dot{x}=\rho x\left( 1-\frac{x}{\kappa}\right)-\frac{\beta x^2}{\alpha^2+ x^2}, $$
où les paramètres $\alpha,\beta,\kappa,\rho$ sont des paramètres positifs. En l’absence de prédateurs, la densité de population des tordeuses $x(t)$ est supposée croître de manière logistique avec un taux de croissance $\rho$ et une capacité de charge $\kappa$. La capacité de charge dépend de la quantité de feuillage restant sur les arbres, mais nous la considérons ici comme fixe, comme mentionné ci-dessus. Le terme $\frac{\beta x^2}{\alpha^2+ x^2}$ représente le taux de mortalité dû à la prédation, principalement par les oiseaux.
La forme spécifique du taux de mortalité prend en compte, de la manière la plus simple, les comportements suivants. Il n’y a presque pas de prédation lorsque les tordeuses sont rares ; les oiseaux cherchent leur nourriture ailleurs. Mais, lorsque la densité de population dépasse un certain niveau critique $x=\alpha$, la prédation se déclenche brusquement puis sature.
On peut maintenant se demander ce que l’on entend par épidémie. Avant de répondre à cette question, il est pratique de mettre le modèle sous une forme adimensionnée, comme nous l’avons fait précédemment pour des modèles plus simples. En effet, le modèle comporte quatre paramètres (positifs) : $\rho,\kappa,\alpha$ et $\beta$. Si vous commencez à jouer le jeu de la remise à l’échelle de l’équation, il est clair qu’il existe plusieurs façons de le faire. Il s’avère, après quelques essais et erreurs, qu’un choix très pratique consiste à se débarrasser des paramètres dans le terme de prédation. Après quelques calculs, on obtient le modèle adimensionnée suivant
$$ \dot{x}=rx\left(1-\frac{x}{K}\right) - \frac{x^2}{1+x^2}\, \cdot $$
Analyse des points fixes.
On a toujours le point fixe $x=0$ et il n’est pas difficile de vérifier qu’il est toujours instable en appliquant le test de la dérivée. Intuitivement, cela n’est pas surprenant puisque pour de très petits $x$, on a approximativement $\dot{x}\approx rx$. Les autres points fixes possibles de l’équation sont donnés par les solutions de l’équation
$$ g(x)=h(x) $$
où$$ g(x)=r\left(1-\frac{x}{K}\right)\quad \text{et}\quad h(x)=\frac{x}{1+x^2}\, \cdot $$
Remarquez que la fonction $h$ est indépendante des paramètres ; seule la fonction $g$, dont le graphe est une ligne droite, change lorsque l’on fait varier $r$ et $K$. Cela provient de la manière dont le modèle a été adimensionné.
Comme le montrent les expériences numériques, si $K$ est suffisamment petit, il y a exactement une intersection pour tout $r>0$. Mais, pour de grands $K$, nous pouvons avoir une, deux ou trois intersections, selon la valeur de $r$. Nous pouvons observer que, lorsque nous sommes dans la gamme des paramètres pour lesquels il y a trois intersections, lorsque nous diminuons $r$ avec $K$ fixe, les points fixes $b$ et $c$ se rapprochent l’un de l’autre et finissent par coïncider. C’est la signature d’une bifurcation col-nœud. Après la bifurcation, il ne reste qu’un seul point fixe (en plus de $0$, bien sûr). Une autre façon de faire varier les paramètres est de fixer $K$ et, en commençant pour un petit $r$, on observe que $a$ et $b$ peuvent « entrer en collision » et s’ « annihiler » lorsque $r$ est augmenté.
Les expériences numériques montrent également comment la stabilité des points fixes change lorsque l’on fait varier $r$ et $K$. Pour les valeurs correspondant à trois points fixes, le plus petit point fixe stable $b$ est appelé le niveau de refuge de la population de tordeuses, tandis que le plus grand point fixe stable $c$ est le niveau d’éclosion. Du point de vue de la lutte contre les parasites, on souhaite maintenir la densité de la population à $a$ et loin de $c$. Le sort du système est déterminé par la condition initiale $x_0$. Une épidémie se produit si et seulement si $x_0>b$ ; le point fixe instable $b$ joue le rôle d’un seuil.
Un autre scénario plus intéressant conduisant à une flambée est celui d’une bifurcation de type col-nœud. Supposons que les paramètres évoluent de manière à faire disparaître le point fixe $a$ : la population passera alors brutalement au niveau épidémique $c$. Et la situation est pire encore, car il existe un effet d’hystérésis : même si l’on rétablit les paramètres initiaux, la population ne redescendra pas au niveau de refuge $a$.
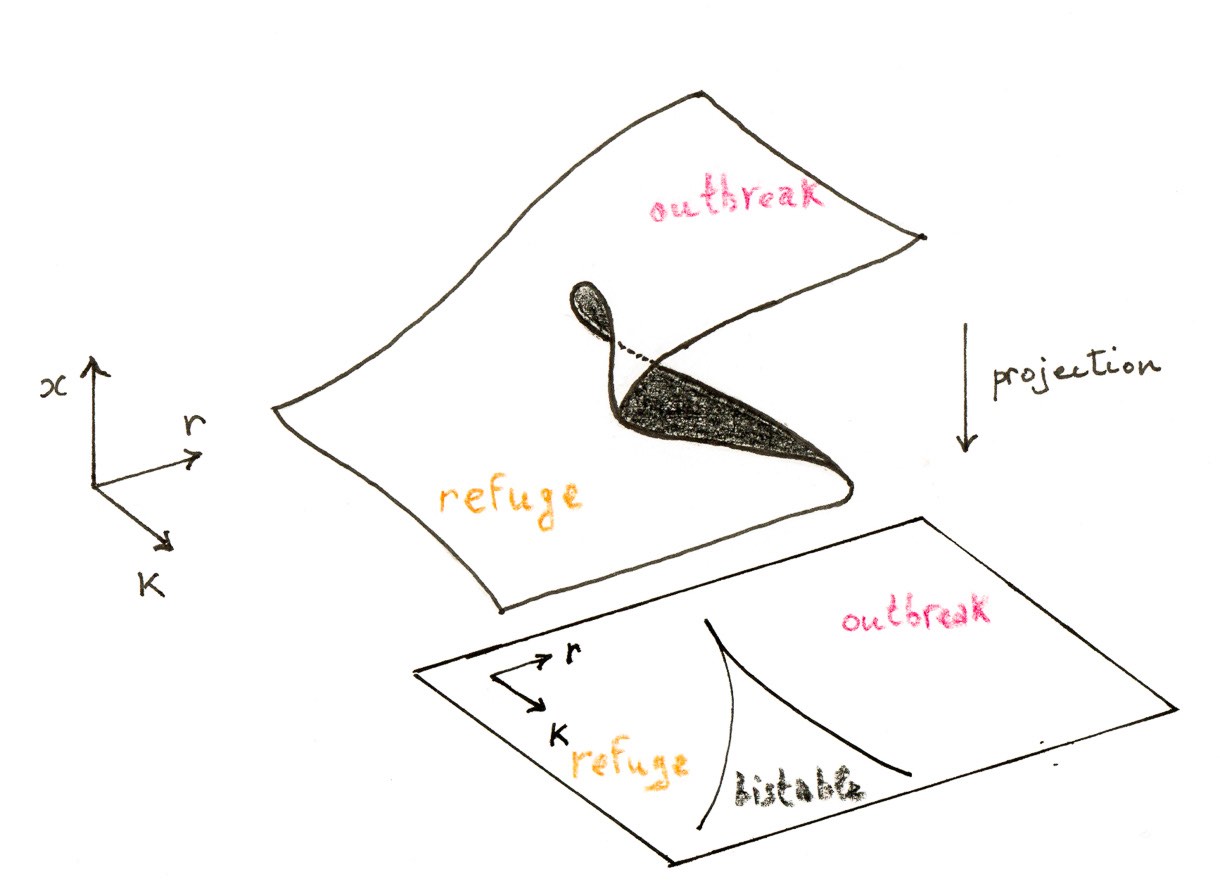
La richesse de ce modèle simple est tout à fait remarquable. Il illustre le fait qu’il ne faut pas sous-estimer les modèles unidimensionnels lorsque des bifurcations peuvent se produire.